
27 May 2020 news
The possible impacts of ocean acidification (OA) on Arctic marine ecosystems is a primary concern, especially on time scales beyond the next election, or beyond our own lifetimes. The Fram Centre’s Ocean Acidification Flagship seeks answers to this challenge.
1.
The cold Arctic Ocean and marginal seas are changing, with higher sea-surface temperatures and less sea ice, but are also getting more acidic (i.e. less basic) as they absorb anthropogenic CO2 from the atmosphere. The CO2 absorbed raises the partial pressure of CO2 (pCO2), increases the hydrogen ion concentration (lowering pH), and removes carbonate ions, thus lowering saturation states for calcium carbonate (Ω).
In a short-term perspective of seasons and years, changes in oceanic pH may not be very noticeable, but in the marine environment and in experiments, the effects of OA are measurable and can potentially become severe for marine organisms that utilize calcium carbonate in their skeletal structures. Non-calcifying organisms can also be affected by OA due to physiological responses that can influence feeding or the costs of maintaining homeostasis, which modulates energetic trade-offs controlling growth and reproduction.
The Ocean Acidification Flagship at the Fram Centre has conducted research projects on different aspects of OA during the last decade.A nine-year oceanographic time series has been established to track the OA state in Fram Strait, investigating both the Arctic outflow waters in the East Greenland Current and in the Atlantic water inflow.
The time series is now starting to show decreased pH and increased pCO2 throughout the water column in the East Greenland Current (Fig. 1). In addition to the Fram Strait study, we investigate the effect of freshwater from glacial melt in Svalbard fjords and have found reductions in alkalinity near the glaciers, with potential consequences for calcifying organisms (Fig. 2).
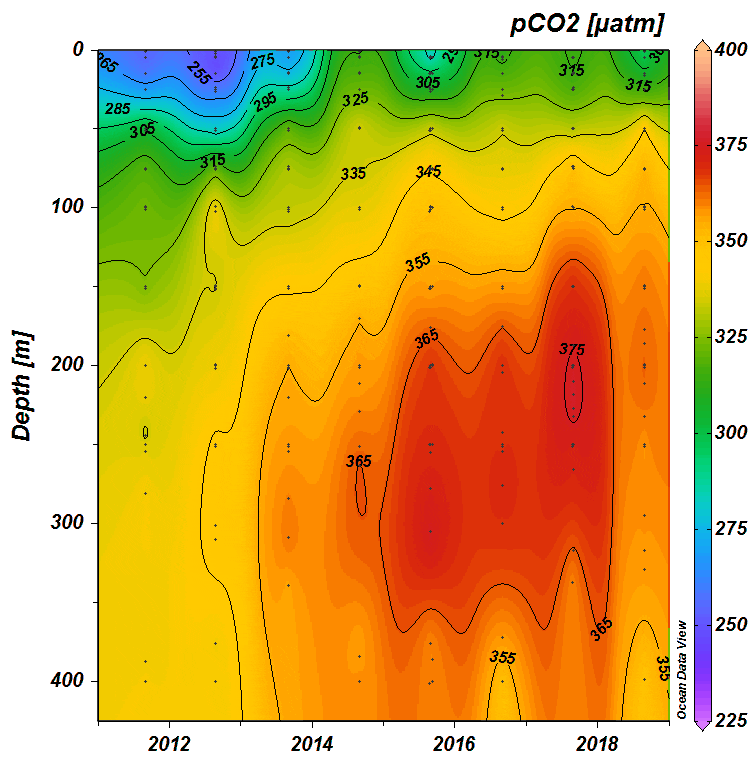 Figure 1. Changes in pCO2 (scale in μatm) in Arctic water exiting the Arctic Ocean in the East Greenland Current from 2011-2018 Graph: M Chierici and A Fransson, unpublished data.
Figure 1. Changes in pCO2 (scale in μatm) in Arctic water exiting the Arctic Ocean in the East Greenland Current from 2011-2018 Graph: M Chierici and A Fransson, unpublished data.
Biological effects
The second part of our flagship deals with biological effects. Studies on calcified skeletons and shells of marine organisms have shown that those with a high proportion of the calcite form of calcium carbonate are less affected than those with more of the aragonite form. Copepods, which have a tough exoskeleton composed of chitin, protein, and calcium carbonate, have important functions in the marine food web. If they were to be affected by OA, the consequences for the pelagic ecosystem would be severe. The good news for both boreal and Arctic copepods (Calanus finmarchicus and C. glacialis) is that they are largely robust to OA, although some life stages and populations are more sensitive than others. Invertebrate DNA may also be vulnerable to ocean acidification-induced damage, and transcriptional changes in DNA repair mechanisms have been reported for Calanus glacialis. We compare the susceptibility of common Arctic copepods (Acartia sp. and Calanus sp.) to DNA damage against that of temperate taxa to estimate the impacts of future low pH on the DNA integrity of these important food-web elements.
 Figure 2. Agneta Fransson collects water sample for carbonate chemistry in Kongsfjorden, Svalbard (Photo: Helene Hodal Lødeme / Institute of Marine Research)
Figure 2. Agneta Fransson collects water sample for carbonate chemistry in Kongsfjorden, Svalbard (Photo: Helene Hodal Lødeme / Institute of Marine Research)  The pteropod sea butterfly Limacina helicina, with a delicate shell of aragonite.
The pteropod sea butterfly Limacina helicina, with a delicate shell of aragonite. 2.
Finding good model organisms for effect studies of OA is challenging. The sea butterfly snail Limacina helicina has been sampled along chemical and physical gradients and used as a proxy for climate-change effects on its shell condition (Fig. 3). Pilot experiments have been done with cladocerans (small crustaceans), whose clonal reproduction allows for investigations into how specific genotypes respond to OA. They have fast life cycles (i.e. weeks), which makes it possible to study them over several generations. Thus, a multigeneration study on the combined effects of temperature and CO2 on gene expression and epigenetic response in Penilia avirostris is currently being performed in order to assess whether it can keep up with the rate of change caused by OA (Fig. 4). Ocean acidification has the potential to affect sexual reproduction and early life-history stages.
The cold-water coral Lophelia pertusa has a long life cycle where embryonic development slows down at higher pCO2 levels, while increased temperature speeds it up.
Long-term monitoring of organisms in field situations to determine the effects of OA has started, but another approach is to use natural analogs to investigate the effects of both climate change and ocean acidification.
Spatial differences in environmental conditions (temperature, salinity, pCO2) can be used as an analog for environmental changes over time. A recent study in Kongsfjorden, Svalbard, sampled shallow-water amphipods (Gammarus setosus) along salinity gradients to determine how different populations respond to salinity as well as elevated pCO2 (Figs. 5-7).
In this case, populations inhabiting low-salinity waters near the glacier front appear to have gained tolerance to the challenge of low salinity, indicating that they may be able to respond similarly to future freshening associated with climate change. The project showed that their physiology responded more to salinity than to elevated pCO2, both of which influence pH.
 Annual mean change in pH (last decade – first decade of simulation) for the NorESM1-ME (left panel) and NORWECOM (middle panel) model. Right panel: Annual mean (0–10 m) pH for Barents Sea (black), the Greenland Sea (red), and the Norwegian Sea (green), for NorESM1-ME (solid line) and NORWECOM (dashed line) (Reproduced with permission from Skogen et al (2018) ICES Journal of Marine Science 75: 2355-2369. https://doi.org/10.1093/icesjms/fsy088)
Annual mean change in pH (last decade – first decade of simulation) for the NorESM1-ME (left panel) and NORWECOM (middle panel) model. Right panel: Annual mean (0–10 m) pH for Barents Sea (black), the Greenland Sea (red), and the Norwegian Sea (green), for NorESM1-ME (solid line) and NORWECOM (dashed line) (Reproduced with permission from Skogen et al (2018) ICES Journal of Marine Science 75: 2355-2369. https://doi.org/10.1093/icesjms/fsy088) 3.
To study the future acidification, as changes in pH, of northern waters and its impacts on marine ecosystems in combination with other stressors, we are using a multi-model approach (Fig. 8). These models are forced by IPCC atmospheric CO2 scenarios to determine basin-wide, regional and local (site-specific) scenarios of biogeochemical and ecosystem change. Recent models have obtained better representations of sea ice, organic carbon, and primary production, as well as improved horizontal resolution. Modelling has also been used to project ecosystem response and feedback to OA, such as potential changes in benthic habitats and communities. How real-world marine food webs absorb change, recover, and adapt (i.e. ecological resilience) to climate change remains a big question. In a modelling project involving long-term data (2004-2016) from Kongsfjorden, we found that the core ecological processes were maintained despite significant environmental perturbations, including ocean acidification. The study showed that Arctic marine food webs can absorb and begin to adapt to ongoing climate change.
 The cladoceran Pernillia avirostris (size 1 mm). (Photo: Albert Lleal / Minden Pictures).
The cladoceran Pernillia avirostris (size 1 mm). (Photo: Albert Lleal / Minden Pictures).
Social science component
The social science component of the Flagship’s work seeks to determine how scientists and environmental managers view, handle, and communicate uncertainties related to OA. We found that scientists have a clear understanding of uncertainty as an integral part of conducting, analysing, and communicating their research.
Environmental managers handle scientific knowledge in the context of multiple stressors and broader ocean management and therefore need ocean acidification knowledge tailored to their reality. The study shows that the differences between scientists and managers should be acknowledged to improve research communication and potential management strategies.
 Allison Bailey with a gammarid amphipod Gammarus setosus. Photo: Allison Bailey / Norwegian Polar Institute
Allison Bailey with a gammarid amphipod Gammarus setosus. Photo: Allison Bailey / Norwegian Polar Institute  Weight-specific oxygen consumption rates for Gammarus setosus can be calculated from the drop in oxygen saturation over time in amphipod respiration chambers. James Brown and Alison Bailey are setting up the experiment on the beach in front of the Marine Laboratory in Ny-Ålesund, with supervision and cheering from Sam Rastrick. (Photo: Wojtek Moskal / Norwegian Polar Institute)
Weight-specific oxygen consumption rates for Gammarus setosus can be calculated from the drop in oxygen saturation over time in amphipod respiration chambers. James Brown and Alison Bailey are setting up the experiment on the beach in front of the Marine Laboratory in Ny-Ålesund, with supervision and cheering from Sam Rastrick. (Photo: Wojtek Moskal / Norwegian Polar Institute)By: Haakon Hop, Agneta Fransson and Allison Bailey // Norwegian Polar Institute. Melissa Chierici, Solfrid Sætre Hjøllo and Sam SP Rastrick // Institute of Marine Research. Philip Wallhead and Marianne Karlsson // Norwegian Institute for Water Research, Bergen. Howard I Browman // Institute of Marine Research, Austevoll Research Station. Claudia Halsband // Akvaplan-niva. Johanna Järnegren // Norwegian Institute for Nature Research, Trondheim
This article was produced for Fram Forum 2020. Fram Forum is published by Framsenteret AS on behalf of FRAM – the High North Research Centre for Climate and the Environment.
